AtaGenix Laboratories

AtaGenix Laboratories
Release time: 2026-06-12 View volume: 284
The cell cycle is the tightly coordinated program governing cell growth, DNA replication, and division, and it lies at the core of proliferation in all cellular life. In eukaryotes, this program is driven by the coordinated activity of cyclins and cyclin-dependent kinases (CDKs), together with quality-control checkpoints at the G1, G2/M, and spindle (M phase) transitions.
The Sulfolobales, an order of thermoacidophilic archaea, serve as a model system for studying the archaeal cell cycle. Their cell cycle is similarly divided into G1, S, G2, M, and D phases, and their division machinery and chromosome segregation mechanisms show striking parallels to those of eukaryotes. Yet until now, the transcriptional regulatory logic underlying cell cycle phase transitions in Sulfolobales had remained largely unresolved. Researchers from the CRISPR and Archaeal Biology Research Center at Shandong University, publishing in Nucleic Acids Research, have now systematically uncovered a cell cycle regulatory mechanism centered on a "wave of repression and de-repression."
The researchers first examined the protein family related to aCcr1, a transcriptional regulator of the cell cycle identified in earlier work. The genome of Saccharolobus islandicus REY15A encodes five ribbon-helix-helix (RHH) domain transcription factors: aCcr1, aCcr2 (SiRe_1157), aCcr3 (SiRe_1577), SiRe_1373, and SiRe_0131.
Using RT-qPCR to track transcript levels across the cell cycle in synchronized cells, the team found that aCcr1 and aCcr3 display clear periodic expression patterns: aCcr1 protein levels peak during cytokinesis (the M/D phase), while aCcr3 peaks at the G1-S transition, with the aCcr1 peak preceding aCcr3's by roughly one hour. By contrast, aCcr2 and the remaining homologs are expressed constitutively throughout the cell cycle. Western blot analysis confirmed this timing pattern at the protein level, with aCcr1 running unusually high on the gel, suggesting it carries a post-translational modification. Together, these findings establish the basic division of labor among the three aCcr proteins: aCcr2 acts as a constitutively expressed, genome-wide repressor, while aCcr1 and aCcr3 carry out stage-specific regulatory roles at defined points in the cell cycle.
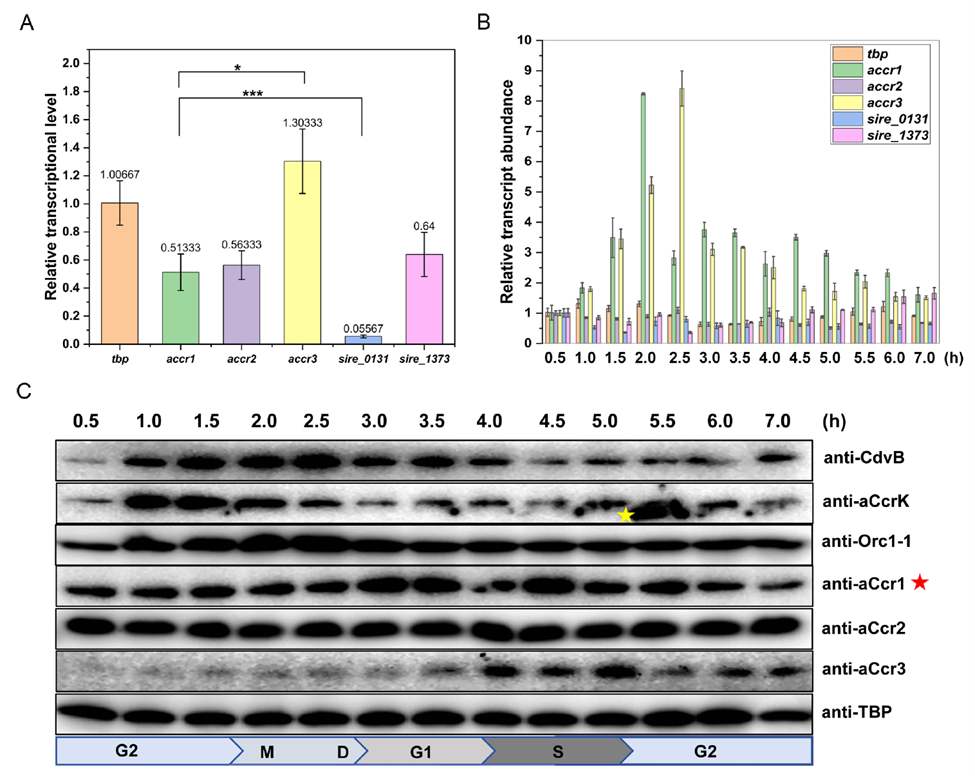
To pin down the functions of aCcr2 and aCcr3 in cell cycle regulation, the researchers built arabinose-inducible overexpression strains in Saccharolobus islandicus. Overexpressing either aCcr2 or aCcr3 produced clear growth retardation, a marked increase in cell size, and elevated genome copy number, closely mirroring the phenotype previously reported for aCcr1 overexpression.
However, subtle differences emerged among the three: aCcr2 overexpression caused the most severe growth inhibition, with aCcr3 falling in between. Flow cytometry showed that, 24 hours after induction, aCcr3-overexpressing cells were predominantly arrested with 2C DNA content, indicating a block prior to G2/M, whereas aCcr1- and aCcr2-overexpressing cells were predominantly polyploid, indicating a block in cell division rather than genome replication. This phenotypic distinction points to differences in the regulatory targets of the three aCcr proteins.
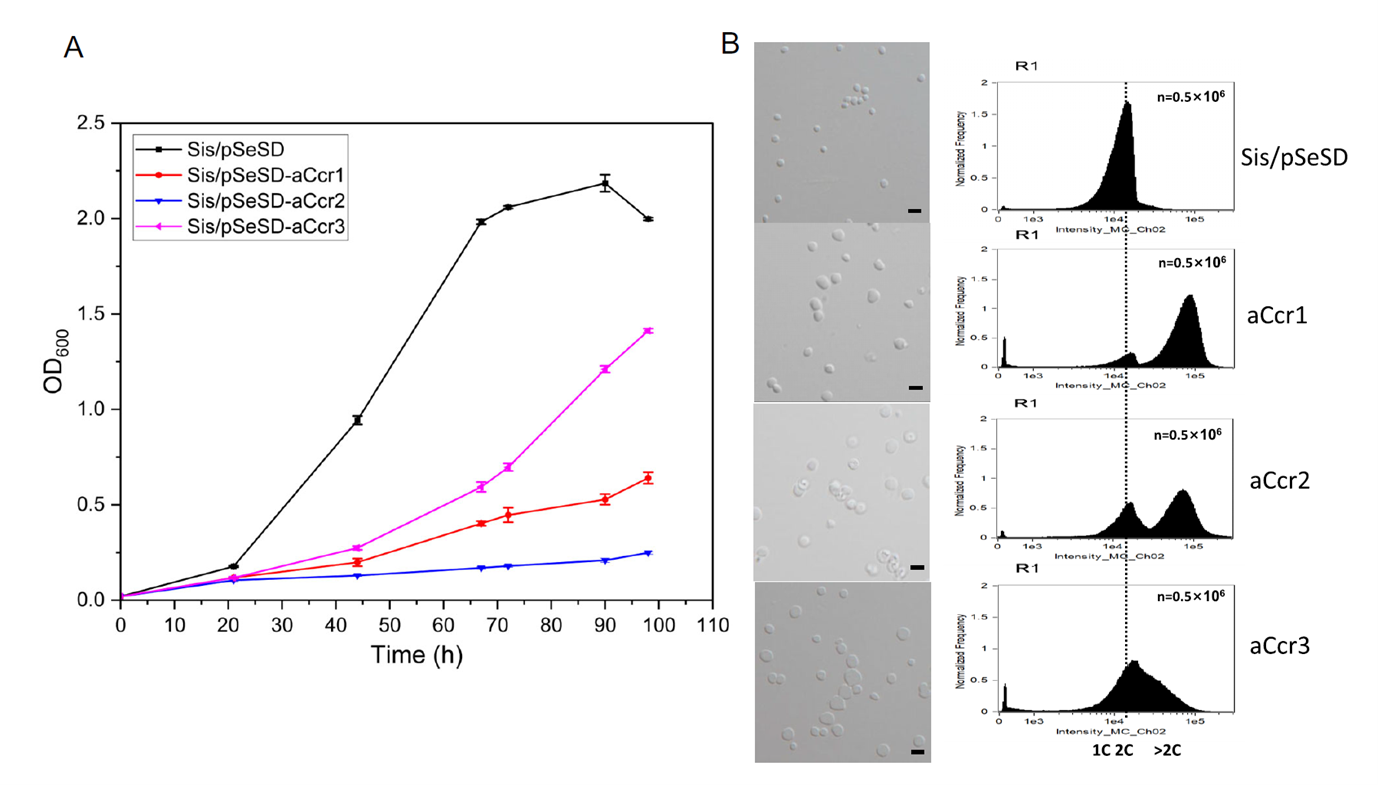
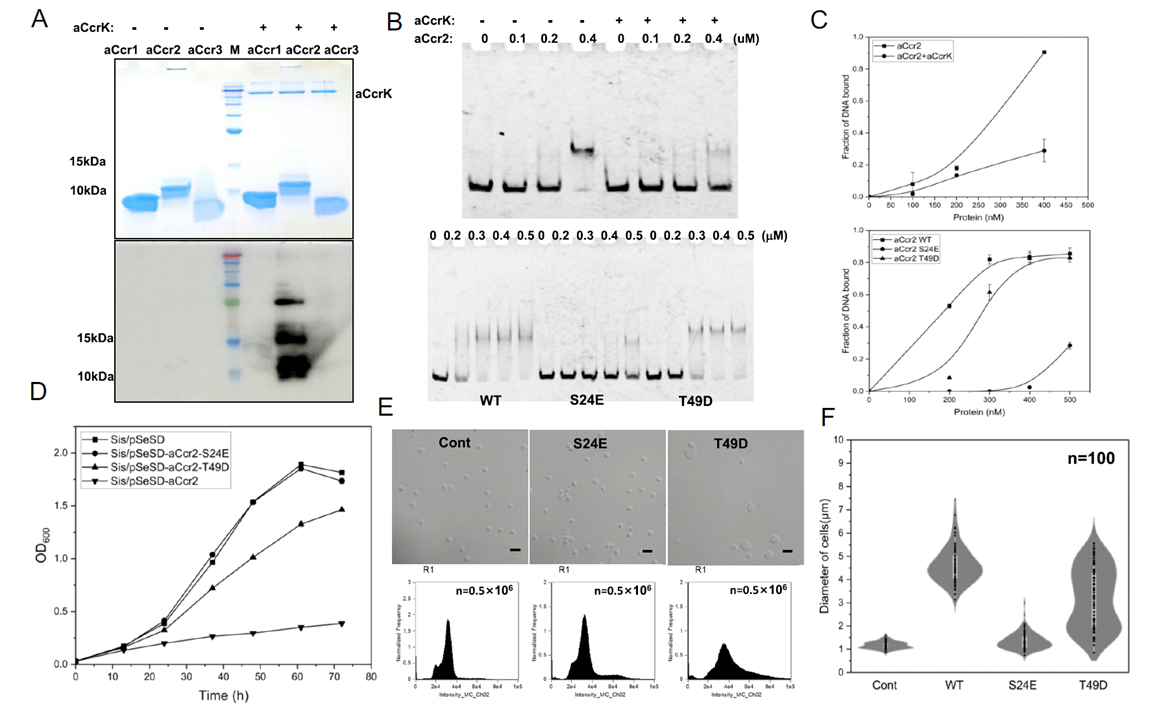
The researchers then used electrophoretic mobility shift assays (EMSA) to characterize the in vitro binding properties of aCcr1, aCcr2, and aCcr3 toward the promoter of cdvA, a gene that initiates cell division. All three proteins bound the cdvA promoter specifically, but with markedly different dissociation constants (KD): aCcr3 showed the highest affinity, aCcr1 an intermediate affinity, and aCcr2 the lowest. This gradient of affinities suggests that once aCcr2 loses its DNA-binding activity through phosphorylation, the higher-affinity aCcr1 or aCcr3 can competitively occupy the same promoter region and re-establish transcriptional repression, providing an orderly mechanism for re-silencing cell cycle genes. This tiered combination of binding affinities provides the molecular basis for the "repression—de-repression—re-repression" timing scheme.
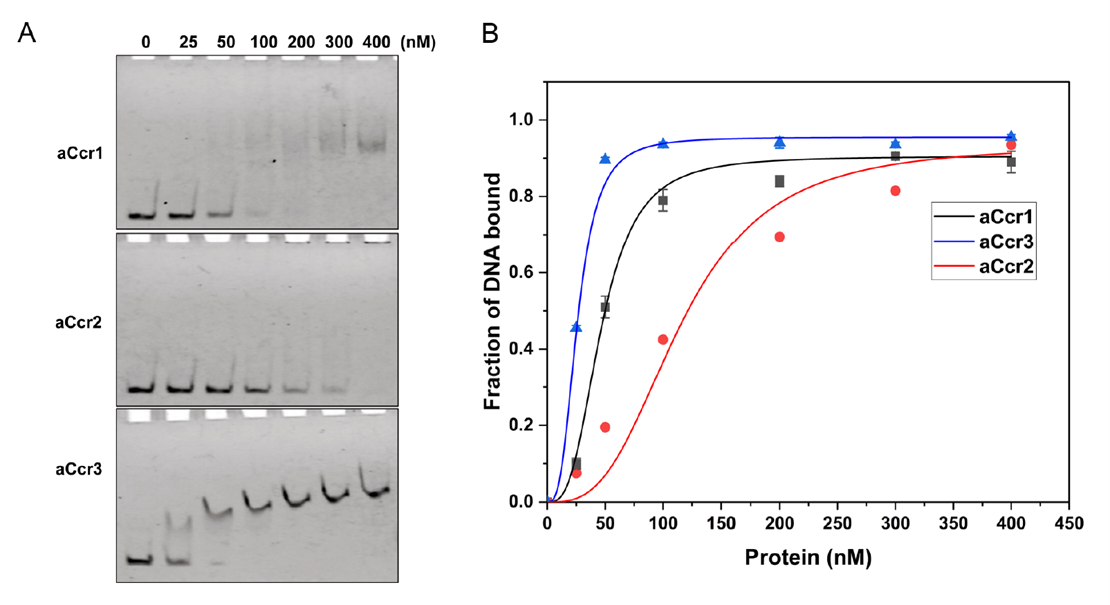
The team carried out genome-wide transcriptomic analyses of the aCcr2- and aCcr3-overexpression strains and compared them systematically with previously published data on aCcr1 overexpression. Overexpressing aCcr2 upregulated 437 genes and downregulated 384; aCcr3 overexpression upregulated 314 genes and downregulated 260; while aCcr1 overexpression affected only 76 upregulated and 124 downregulated genes. Among the 50 genes commonly downregulated by all three, 20% are known essential genes, 48% show periodic expression, and 60% are located in the transcriptionally active chromosomal A region. These 50 genes are largely involved in cell division (cdvA, cdvB, cdvC), chromosome segregation (segA, segB, segC), DNA replication and repair (orc1-1, gins, endoMS), and chromatin organization (cren7).
Further ChIP-seq analysis identified 1,407 genome-wide binding peaks for aCcr2, far exceeding the 224 peaks for aCcr3 and 610 for aCcr1. Comparative analysis suggested that aCcr2's binding motif is more degenerate, which may explain why it regulates the largest number of genes, whereas the more stringent motifs of aCcr1 and aCcr3 correspond to their precise, stage-specific regulatory roles.
The constitutive high-level expression of aCcr2 implies that some mechanism must inactivate it at a specific point in the cell cycle to relieve repression of key genes. The researchers noted that, among synchronized cells, only ePK2 (a eukaryotic-like kinase, now renamed aCcrK) showed clear periodic expression, peaking at the end of G2—precisely the window just before cells enter division.
In vitro experiments confirmed that aCcrK specifically phosphorylates aCcr2, with no comparable effect on aCcr1 or aCcr3. This substrate specificity suggests the phosphorylation event serves a precise regulatory purpose rather than being incidental. Phosphorylated aCcr2 showed markedly reduced DNA-binding ability. Mass spectrometry identified the phosphorylation sites as Ser24 and Thr49, with Ser24 being the most critical—an aCcr2 variant carrying the phosphomimetic mutation S24E almost completely lost its ability to bind the promoter and no longer caused growth inhibition or morphological abnormalities in cells. Thr49 had a more modest effect, and the effect of Thr6 phosphorylation was independently validated through structural modeling and mutagenesis.
These results indicate that aCcrK-mediated phosphorylation of aCcr2 is the core molecular mechanism that relieves transcriptional repression of key genes and licenses cell cycle progression just before cells enter M phase.
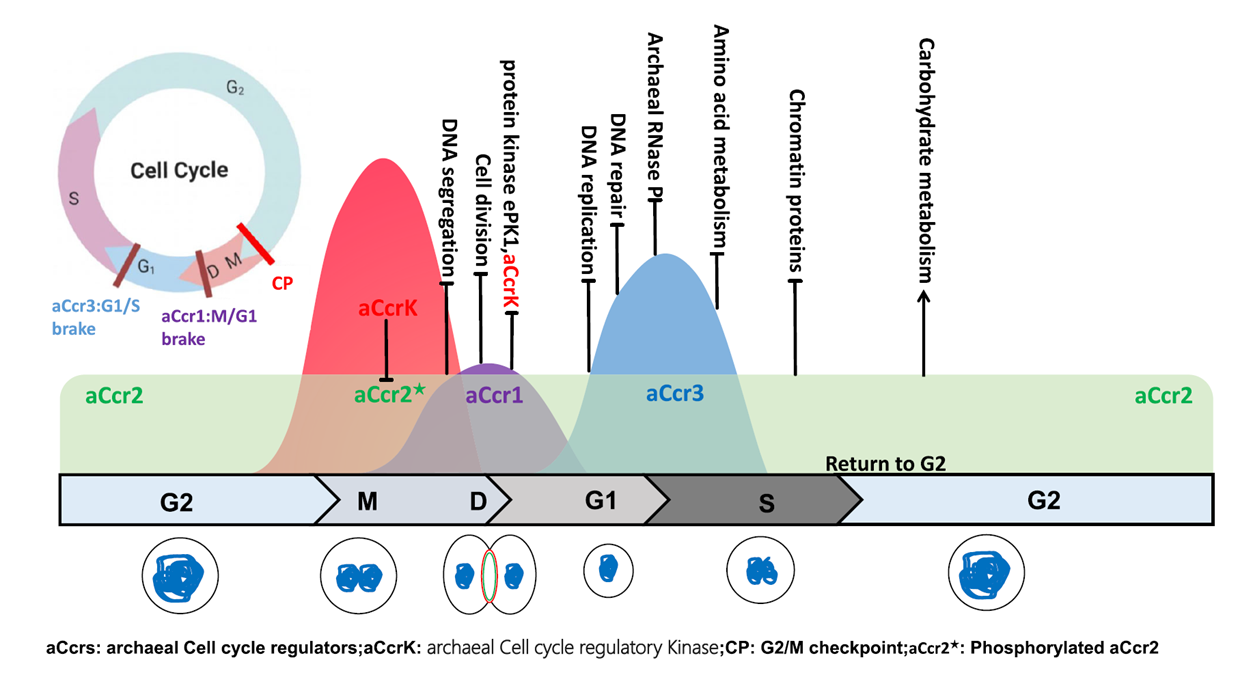
The cell cycle of Sulfolobales archaea shows structural phases strikingly similar to those of eukaryotic cells, yet the underlying transcriptional regulatory mechanism had long remained unclear. This study systematically reveals the cooperative regulatory logic of three RHH-domain transcription factors—aCcr1, aCcr2, and aCcr3—in the cell cycle of Saccharolobus islandicus: aCcr2, expressed throughout the cell cycle, acts as a broad-spectrum repressor; phosphorylation by aCcrK at the end of G2 relieves its repression of key cell cycle genes, opening a window for cells to enter M phase. Subsequently, the periodically upregulated aCcr1 and aCcr3, with higher binding affinities, re-occupy the same promoters, establishing "braking points" at the M/G1 and G1/S transitions, respectively, and keeping the orderly progression of the cell cycle on a controlled track.
This "wave of repression—phosphorylation-driven de-repression—re-repression" regulatory logic does not rely on transcriptional activation, but instead is driven by the precise temporal control of transcriptional repression—representing what may be a simplified intermediate form along the evolutionary path toward eukaryotic cell cycle checkpoint regulation.
For this study, AtaGenix custom-produced polyclonal antibodies against aCcr2, aCcrK, and aCcr3 for the research team. Using these antibodies, the team confirmed the distinct expression timing of the three proteins across the cell cycle by Western blot, and mapped the genome-wide chromatin binding profiles of aCcr2 and aCcr3 by ChIP-seq—the former providing protein-level evidence for the "wave of repression" model, and the latter directly defining the target gene sets and functional boundaries of each regulator.
AtaGenix specializes in custom antibody development with a focus on high specificity
Our antibodies have supported numerous high-impact mechanistic studies across Western blot, ChIP-seq, IF, and other applications
Contact us today to develop your custom research antibody
Contact Us
+86-27-6552-3339
info@atagenix.com
Building C, R & D Building, No. 666, Shendun 4th Road, Donghu New Technology Development Zone, Wuhan
